Duranti S1, Mancabelli L1, Mancino W1, Anzalone R2, Longhi G2, Statello R3, Carnevali L3, Sgoifo A3, Bernasconi S4, Turroni F1, Ventura M1
1. Phòng thí nghiệm Probiogenomics, Khoa Hóa học, Khoa học Đời sống và Tính bền vững Môi trường, Đại học Parma, Parma, Ý.
2. GenProbio srl Parma, Parma, Ý.
3. Phòng thí nghiệm Sinh lý học căng thẳng, Khoa Hóa học, Khoa học Đời sống và Môi trường Bền vững, Đại học Parma, Parma, Ý.
4. Trung tâm nghiên cứu Microbiome, Đại học Parma, Parma, Ý.
Tóm tắt nội dung
COLOSTRONONI là một thực phẩm chức năng bao gồm sữa non của bò và quả Morinda citrifolia (Noni). Trong nghiên cứu này, chúng tôi đã kiểm tra khả năng của COLOSTRONONI trong việc ảnh hưởng đến thành phần hệ vi sinh vật đường ruột bằng cách sử dụng đánh giá in vivo ở chuột. Hơn nữa, chúng tôi đã phân tích tác động của COLOSTRONONI đối với các phản ứng viêm toàn thân cũng như tính thấm ruột của động vật. Nhìn chung, các phân tích của chúng tôi đã ủng hộ khái niệm COLOSTRONONI như một loại thực phẩm bổ sung tự nhiên không ảnh hưởng (không tiêu cực và tích cực) đến cân bằng nội môi hệ vi sinh vật đường ruột trong điều kiện khỏe mạnh. Hơn nữa, COLOSTRONONI nhấn mạnh tác dụng thấp hơn trong việc biểu hiện các gen mã hóa cho phản ứng IL-10, Il-12 và TNF-α cho phép chúng tôi đưa ra giả thuyết về hoạt động điều hòa miễn dịch của thực phẩm chức năng này.
Giới thiệu
COLOSTRONONI là một thực phẩm bổ sung chế độ ăn uống mới bao gồm sữa non của bò và quả Morinda citrifolia (Noni) [ 1 ]. Một số bằng chứng khoa học cho thấy rằng sữa non của bò và trái nhàu có tác dụng điều hòa ngăn ngừa tình trạng viêm đường ruột và do đó, có thể làm giảm sự phát triển của các bệnh toàn thân mãn tính do viêm [ 1 ].
Sữa non là sữa đầu tiên được lấy từ tuyến vú cho con bú sau khi sinh. Nó thể hiện một khía cạnh độc đáo trong dinh dưỡng của trẻ sơ sinh, trên thực tế, nó có thể thúc đẩy sự phát triển của thành phần vi sinh vật của đường tiêu hóa cũng như hệ thống miễn dịch thông qua việc thu nhận các globulin miễn dịch của mẹ [ 2 ]. Sữa non của bò rất giàu các cytokine gây viêm bao gồm interleukin (IL-2, IL-1β, IL-6, IL-17) và các globulin miễn dịch, chẳng hạn như IgA, IgM và IgG, yếu tố hoại tử khối u-α (TNF-α), interferon- γ, và các hợp chất không kháng khuẩn khác góp phần bảo vệ sinh vật chống lại sự lây nhiễm vi sinh vật gây bệnh [ 3 , 4]. Ngoài ra, sữa non của bò bao gồm các thành phần liên quan đến hệ thống miễn dịch bẩm sinh như peptide kháng khuẩn cũng như lactoferrin và lactoperoxidase, thể hiện các tác dụng kháng khuẩn bổ sung [ 4 , 5 ].
Các Morinda citrifolia L., còn được gọi là Noni, là một nhà máy sản xuất hơn 160 chất phytochemical được xác định, trong đó có nguồn gốc ở Ấn Độ Dương và Polynesia và nó được phân phối rộng rãi trên toàn thế giới [ 6 ]. Các hợp chất sinh học như glycoside, polysaccharide, trisaccharide axit béo este, vitamin và khoáng chất đã được phân lập từ quả, rễ và lá cây nhàu [ 7 ]. Các nghiên cứu gần đây tiết lộ rằng nước trái cây đã chiết xuất từ M . Quả citrifolia (Noni) có một loạt các tác dụng điều hòa miễn dịch, bao gồm các hoạt động kháng khuẩn, chống viêm, chống tạo khối u và chống oxy hóa [ 7 – 11 ].
Hàng rào ruột có khả năng điều chỉnh sự hấp thụ chất dinh dưỡng và sự trao đổi các phân tử giữa vật chủ và môi trường [ 12 ]. Cụ thể, các điểm nối chặt chẽ giữa các nội mạc (TJ) đóng một vai trò quan trọng trong tính thấm của hàng rào nội mô [ 13 ]. Đặc biệt, các TJ kết nối các tế bào lân cận với nhau để tạo ra một hàng rào ngăn chặn sự lây nhiễm mầm bệnh cũng như điều chỉnh sự khuếch tán nội bào của các ion và chất hòa tan [ 14 ].
Mục đích của nghiên cứu này là đánh giá ảnh hưởng của COLOSTRONONI đối với đa dạng sinh học vi sinh vật tổng thể trong đường ruột của chuột, nhằm xác định tác động giả định của thực phẩm bổ sung này đối với các nhóm vi khuẩn cụ thể. Hơn nữa, trong nghiên cứu này, chúng tôi đánh giá tác động của COLOSTRONONI đối với các phản ứng viêm hệ thống cũng như tính thấm ruột của chuột.
Vật liệu và phương pháp
Tuyên bố đạo đức
Tất cả các quy trình và quy trình thử nghiệm liên quan đến động vật đã được Ủy ban chăm sóc và sử dụng động vật bác sĩ thú y của Đại học Parma (số 370/2018) phê duyệt và được tiến hành theo Chỉ thị của Hội đồng Cộng đồng Châu Âu ngày 22 tháng 9 năm 2010 (2010/63 / UE).
Chuồng thú
Các thí nghiệm liên quan đến chuột Groningen loại hoang dã đực ba tháng tuổi ( Rattus norvegicus ), ban đầu được lấy từ Đại học Groningen (Hà Lan), và được nhân giống trong các cơ sở động vật ở điều kiện tiêu chuẩn. Sau khi cai sữa, những con chuột được nuôi trong các nhóm anh chị em cùng giới tính trong phòng trong điều kiện kiểm soát độ ẩm – (50 ± 10%) và nhiệt độ (22 ± 2 ° C), chu kỳ sáng – tối 12 giờ (đèn bật sáng lúc 7 giờ sáng), và với thức ăn và nước uống có sẵn ad libitum .
Mô tả COLOSTRONONI
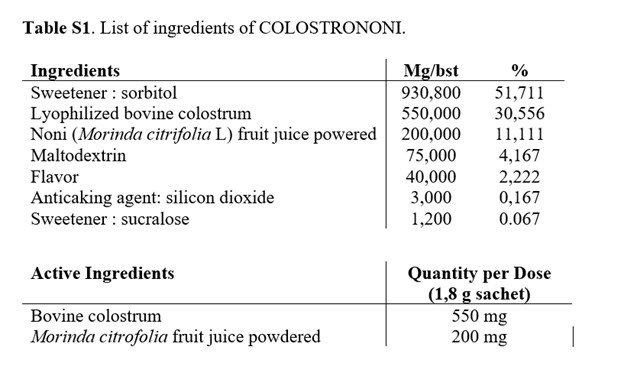
COLOSTRONONI là một loại thực phẩm chức năng bao gồm 30,56% sữa non bò và 11,11% nước ép trái cây Morinda citrifolia (Noni) ( Bảng S1 ). Bảng quy trình sản xuất (Colostrononi 2205C02) được hiển thị trong Hình S1 . Danh sách tất cả các thành phần được mô tả trong Bảng S1 . Hơn nữa, việc kiểm tra chất lượng được thực hiện bởi Phòng thí nghiệm Verdellino, Ý và số chứng chỉ phân tích là 201727122. Các chi tiết liên quan đến việc kiểm tra chất lượng được mô tả trong Bảng S2 . Các thí nghiệm về độ ổn định cho thấy thời hạn sử dụng của COLOSTRONONI là 36 tháng ( Bảng S3 ). Hơn nữa, sản phẩm được bảo quản nơi khô ráo thoáng mát, tránh ánh nắng trực tiếp.
Cuối cùng, các đặc tính hóa học của Sữa non bò và nước trái cây Morinda citrifolia được cung cấp trong Bảng S4 và S5 .




Thiết kế thử nghiệm của các thử nghiệm in vivo
Tiến trình của tất cả các thủ tục được minh họa trong Hình 1. Cụ thể, những con chuột được nhốt riêng lẻ trong lồng polymethyl methacrylate (Plexiglas) (39 cm × 23 cm × 15 cm). Trong tuần đầu tiên, động vật tiếp tục theo chế độ ăn chow bình thường nhưng được uống hàng ngày dung dịch sucrose 2% (thể tích: 500μl), để huấn luyện chúng uống tự phát từ ống tiêm. Sau đó, chuột được phân ngẫu nhiên thành hai nhóm (n = 10 mỗi con) và được cho ăn với chế độ ăn tiêu chuẩn có bổ sung COLOSTRONONI (Số sản phẩm 934744602, Lô # A04268, Guna Spa, Milan, Ý) được hòa tan trong dung dịch đường sucrose (2%) ( Nhóm CN) [0,500 gr / kg (w / w)], hoặc duy trì chế độ ăn tiêu chuẩn có bổ sung dung dịch sucrose (2%) (nhóm CTRL) trong 12 ngày tiếp theo. Cuối cùng, vào cuối quy trình thử nghiệm, những con chuột đã được chết. Hơn nữa,Hình 1 ). Tại cùng thời điểm, BW được đo lường và các thay đổi BW được tính toán như đã mô tả trước đây [ 15 ].

Thời gian và kết quả của quy trình thí nghiệm.
Bảng A hiển thị lịch trình của các quy trình thí nghiệm. Bảng B và C hiển thị các thay đổi về trọng lượng cơ thể và tỷ lệ phần trăm ăn vào so với các giá trị ‘T0’ tương ứng, trong quá trình thử nghiệm. Giá trị được biểu thị dưới dạng trung bình ± sai số chuẩn của giá trị trung bình (SEM).
Tách chiết DNA vi khuẩn phân và manh tràng
Các mẫu phân được tách chiết DNA bằng Bộ công cụ nhỏ phân DNA QIAmp theo hướng dẫn của nhà sản xuất (Qiagen). Hơn nữa, chiết xuất DNA vi khuẩn từ các mẫu manh tràng được thực hiện bằng cách sử dụng PowerViral Environmental RNA / DNA Isolation Kit Sample (Qiagen) theo hướng dẫn của giao thức. Nồng độ và độ tinh khiết của DNA sau đó được xác định bằng Máy quang phổ Picodrop microlit (Picodrop).
Khuếch đại và giải trình tự gen 16S rRNA
Một phần trình tự gen 16S rRNA được khuếch đại từ DNA chiết xuất bằng cách sử dụng cặp mồi Probio_Uni và / Probio_Rev, nhắm mục tiêu vào vùng V3 của trình tự gen 16S rRNA [ 16]. Bộ điều hợp Illumina trình tự nucleotide nhô ra được thêm vào amplicon đặc hiệu gen 16S rRNA một phần, được xử lý thêm bằng cách sử dụng Giao thức chuẩn bị thư viện trình tự gen 16S (Phần # 15044223 Rev. B – Illumina; xem thêm bên dưới). Phù hợp với các giao thức Illumina, các sản phẩm PCR thu được sau quá trình khuếch đại trình tự gen 16S rRNA đã được tinh sạch bằng bước tinh sạch từ tính liên quan đến các hạt tinh sạch DNA Agencourt AMPure XP (Beckman Coulter Genomics GmbH, Bernried, Đức) để loại bỏ các đoạn mồi. Nồng độ DNA của thư viện trình tự khuếch đại được ước tính thông qua hệ thống định lượng Flometric Qubit (Life Technologies). Amplicon được pha loãng đến 4 nM và 5 μl của mỗi amplicon DNA pha loãng được trộn để chuẩn bị cho thư viện cuối cùng được gộp chung.
Phân tích cấu hình vi khuẩn rRNA 16S
Theo trình tự, tệp fastq được xử lý bằng tập lệnh tùy chỉnh dựa trên bộ phần mềm QIIME [ 17 ]. Các cặp đầu đọc được ghép nối đã được lắp ráp để tạo lại các amplicon Probio_Uni / Probio_Rev hoàn chỉnh. Kiểm soát chất lượng giữ lại các trình tự có độ dài từ 140 đến 400 bp và điểm chất lượng trình tự trung bình> 20 trong khi các trình tự có homopolyme> 7 bp và mồi không khớp được bỏ qua. Để tính toán các phép đo phân tập xuôi dòng (chỉ số phân tập alpha và beta, phân tích Unifrac), Đơn vị phân loại hoạt động 16S rRNA (OTU) được xác định ở mức tương đồng trình tự ≥ 99% bằng cách sử dụng uclust [ 18 ] và OTU có ít hơn 10 trình tự đã được lọc. Tất cả các lần đọc được phân loại theo thứ hạng phân loại thấp nhất có thể sử dụng QIIME [ 17 ] và tập dữ liệu tham chiếu từ cơ sở dữ liệu SILVA [ 19] cho dữ liệu rRNA 16S. Đa dạng sinh học của các mẫu (đa dạng alpha) được tính toán với các chỉ số Chao1 và Shannon. Sự tương đồng giữa các mẫu (đa dạng beta) được tính bằng uniFrac có trọng số và không có trọng số [ 20 ]. Phạm vi tương đồng được tính toán giữa các giá trị 0 và 1. Các biểu diễn PCoA của phân tập beta được thực hiện bằng QIIME [ 17 ].
Phân tích thống kê
Phân tích PERMANOVA được thực hiện, sử dụng QIIME [ 17 ] để đánh giá sự khác biệt về thành phần vi sinh vật giữa nhóm CN và CTRL. Tất cả dữ liệu được trình bày dưới dạng phương tiện ± SEM So sánh giữa hai nhóm được thực hiện bằng phép thử t của Học sinh chưa được ghép nối. Sự khác biệt quan sát được tương ứng với P <0,05, P <0,01 và P < 0,001 lần lượt được coi là có ý nghĩa thống kê, ý nghĩa cao hoặc cực kỳ cao. Tất cả các phân tích được thực hiện bằng phần mềm Prism 5 (GraphPad Software Inc. San Diego, CA).
Phân lập RNA và biểu hiện gen
RNA tổng số được phân lập bắt đầu từ 1 gr mô caeca được lơ lửng trong 1 ml QIAzoL (Qiagen, Vương quốc Anh) và đặt trong một ống có chứa 0,8 g hạt thủy tinh (đường kính, 106 μm; Sigma). Các tế bào được ly giải bằng cách lắc hỗn hợp trên máy đồng nhất BioSpec ở 4 ° C trong 2 phút (cài đặt tối đa). Sau đó, hỗn hợp này được ly tâm với tốc độ 12.000 vòng / phút trong 15 phút, và giai đoạn trên có chứa mẫu chứa RNA được thu hồi. Mẫu RNA được tinh chế thêm bằng RNeasy Mini Kit (QIAGEN) theo các giao thức cụ thể. Chất lượng của RNA được kiểm tra bằng cách phân tích tính toàn vẹn của các phân tử rRNA bởi Trạm MultiTape (Agilent). Phiên mã ngược sang cDNA được thực hiện với bộ tổng hợp cDNA iScript Select (Phòng thí nghiệm Bio-Rad) theo hướng dẫn của nhà cung cấp. Các mức biểu hiện mRNA được phân tích bằng công nghệ xanh SYBR trong PCR thời gian thực định lượng (qRT-PCR) sử dụng QPCR Green Master Mix LRox (Biotech Rabbit GmbH) trên hệ thống Bio-Rad CFX96 theo hướng dẫn của nhà sản xuất (Bio-Rad). Các loại sơn lót được sử dụng được chỉ định trong S6 Bảng trong tài liệu bổ sung. PCR định lượng được thực hiện theo chu kỳ sau: ban đầu giữ ở 96 ° C trong 30 s và sau đó 42 chu kỳ ở 96 ° C trong 15 s và Tm ° C trong 30 s. Ý nghĩa thống kê giữa các phương tiện được phân tích bằng cách sử dụng thử nghiệm Student t không ghép đôi với ngưỡng P <0,05. Giá trị được biểu thị bằng giá trị trung bình ± SEM của chuột mỗi nhóm. Tính toán thống kê được thực hiện bằng chương trình phần mềm GraphPad Prism 5.

Lắng đọng dữ liệu
Trình tự thô của cấu hình gen 16S rRNA có thể truy cập được thông qua số gia nhập nghiên cứu SRA PRJNA497777.
Kết quả và thảo luận
Sự phát triển của động vật và lượng thức ăn
Để đánh giá xem COLOSTRONONI có thể điều chỉnh các điều kiện sinh lý của chuột hay không, chúng tôi đã thực hiện một thí nghiệm in vivo bằng cách sử dụng hai nhóm chuột, một nhóm nhận COLOSTRONONI (CN) trong chế độ ăn của chúng và nhóm còn lại duy trì theo chế độ ăn tiêu chuẩn (CTRL). Ảnh hưởng của COLOSTRONONI lên trọng lượng cơ thể (BW), cũng như lượng thức ăn (FI) của động vật đã được ghi nhận ( Hình 1 ).
ANOVA hai chiều đối với các phép đo lặp lại mang lại ảnh hưởng đáng kể về thời gian đối với các thay đổi BW ( F = 232,09, p -value <0,001) và FI / BW ( F = 22,40, p -value <0,001). BW tăng tương tự ở cả hai nhóm trong quá trình thử nghiệm. Tương tự, không có sự khác biệt đáng kể giữa hai nhóm được tìm thấy trong các thay đổi FI. Đáng chú ý, FI dường như chỉ tăng trong tuần đầu tiên của điều kiện CN / CTRL (CN: + 3,17%; CTRL: + 4,12%).
Những dữ liệu này cho thấy rằng điều trị COLOSTRONONI không ảnh hưởng đến sự phát triển của BW cũng như FI ở chuột.
Đặc điểm tác động của COLOSTRONONI lên thành phần hệ vi sinh vật đường ruột của chuột
Các mẫu phân và manh tràng được sử dụng để xác định cấu trúc phân loại vi sinh vật có liên quan đến thực phẩm chức năng COLOSTRONONI hay không bằng phương pháp phân tích trình tự rRNA 16S như đã mô tả trước đây [ 16 ]. Trình tự Thế hệ tiếp theo của 80 mẫu, bao gồm cả mẫu phân ở các thời điểm khác nhau và mẫu manh tràng, tạo ra tổng cộng 5.727.788 lần đọc, dao động từ 43.050 đến 92.936 mỗi mẫu ( Bảng S7 ). Chất lượng và lọc chimera dẫn đến tổng số 4.863.323 lần đọc được lọc với mức trung bình là 60.791 và dao động từ 27.534 đến 82.880 lần đọc cho mỗi mẫu ( Bảng S7 ). Các đường cong đa dạng sinh học Chao1 gợi ý rằng trong mọi trường hợp, độ sâu giải trình tự thu được là thích hợp để bao phủ phần lớn đa dạng sinh học của hệ vi sinh vật có trong các mẫu (Hình 2A và 2B ). Đáng chú ý, việc bổ sung CN không liên quan đến sự thay đổi rõ ràng và / hoặc giảm đa dạng sinh học của hệ vi sinh vật đường ruột trong các mẫu manh tràng cũng như trong các mẫu phân trong tất cả các giai đoạn xử lý (p -value> 0,05 được tính toán thông qua thử nghiệm t của Student không ghép đôi ) ( Hình 2C ). Để đánh giá sự khác biệt về hệ vi sinh vật chuột giữa nhóm CN và CTRL, chúng tôi đã phân tích sự đa dạng beta dựa trên UniFrac không trọng số và có trọng số cho các nhóm này, sau đó là Phân tích Tọa độ Chính (PCoA) ( Hình 3 ). Phân tích PCoA được thực hiện trên các mẫu phân tại các thời điểm khác nhau cũng như trên các mẫu manh tràng, riêng biệt. Những dữ liệu này làm nổi bật sự xuất hiện, đối với cả hai nhóm, của chỉ một cụm ( Hình 3), gợi ý duy trì sự ổn định của thành phần hệ vi sinh vật đường ruột. Đáng chú ý, những kết quả này cũng được xác nhận bởi phân tích thống kê PERMANOVA (p-value> 0,05) ( Hình 3 ).


Bảng a và b báo cáo mức độ đa dạng alpha trung bình thu được bằng cách sử dụng chỉ số Chao1 cho các mẫu manh tràng và phân. Bảng c cho thấy hộp và âm mưu râu ria. Các dải bên trong hộp là đường trung bình, các đầu của râu biểu thị mức tối thiểu và tối đa của tất cả dữ liệu.
Bảng a và b bao gồm PCoA minh họa sự đa dạng beta của các mẫu manh tràng CN và CTRL được tính toán thông qua UniFrac không trọng số và có trọng số, tương ứng. Bảng c và d cho thấy trong PCoA báo cáo sự đa dạng beta của các mẫu phân CN và CTRL được tính toán tương ứng thông qua UniFrac có trọng số và có trọng số. Nó cũng đã được báo cáo giá trị p được tính toán thông qua phân tích PERMANOVA.
Thành phần hệ vi sinh vật trong manh tràng của chuột được chứng minh là thống trị bởi Firmicutes phylum (giá trị trung bình 79,92%), tiếp theo là Bacteroidetes (giá trị trung bình 18,07%). Ngược lại, phân tích thành phần hệ vi sinh vật trong phân cho thấy sự phong phú cao của Bacteroidetes (giá trị trung bình 62,27) và Firmicutes phylum (giá trị trung bình 36,61%). Đơn vị phân loại vi khuẩn phong phú nhất là họ nhóm Bacteroidales S24-7 (lần lượt là 15,67% và 54,29% trong mẫu manh tràng và trong mẫu phân) và hai chi khác nhau thuộc họ Lachnospiraceae (tổng số trung bình là 41,00% 18,14% trong mẫu manh tràng và trong phân mẫu tương ứng) (Hình 4 và 5). Việc so sánh hệ vi sinh vật của các mẫu manh tràng và phân của chuột CN và CTRL tương ứng cho thấy không có sự khác biệt đáng kể về thành phần hệ vi sinh vật giữa mỗi nhóm. Những kết quả này ủng hộ giả thuyết rằng sản phẩm COLOSTRONONI không ảnh hưởng (không tiêu cực và tích cực) đến cân bằng nội môi của hệ vi sinh vật đường ruột trong điều kiện khỏe mạnh, duy trì trạng thái cân bằng giữa các đơn vị vi khuẩn khác nhau cấu thành hệ vi sinh vật đường ruột. Tập trung vào chi Akkermansia , tức là Akkermansia muciniphila, được báo cáo là thực hiện các hoạt động tăng cường sức khỏe thông qua việc điều chỉnh hệ thống miễn dịch của vật chủ và bằng cách tăng sinh các tế bào T điều hòa chống viêm, chúng tôi đã quan sát thấy sự gia tăng nhẹ của chi này trong các mẫu phân được xử lý CN (mức trung bình tương đối dồi dào là 0,66 % ± 1,80%, 1,97% ± 4,20% và 1,39% ± 3,19% ở T0, T1 và T2 tương ứng) so với nhóm đối chứng (mức trung bình tương đối 0,02% ± 0,05%, 0,22% ± 0,53% và 0,12% ± 0,30% ở T0, T1 và T2 tương ứng) [ 21 ]. Cụ thể, tỷ lệ phổ biến thu được từ việc so sánh từng mẫu T0 với mẫu T1 hoặc T2 tương ứng trong nhóm CN cho thấy sự gia tăng của đơn vị phân loại này trong 50% trường hợp (6 mẫu T1 và 4 mẫu T2) ( Hình 5). Ngược lại, tỷ lệ so sánh của mỗi mẫu T0 với mẫu T1 hoặc T2 tương ứng trong nhóm CTRL cho thấy sự gia tăng của đơn vị phân loại này chỉ trong 25% trường hợp (3 mẫu T1 và 2 mẫu T2) ( Hình 5 ).

Biểu đồ thanh cho thấy mức trung bình của sự thay đổi mức độ phong phú tương đối ở CN đối với chuột CTRL ở cấp độ chi. Chỉ các đơn vị phân loại có mức độ phong phú tương đối trung bình> 1% trong các nhóm này được hiển thị.

Biểu đồ thanh cho thấy sự khác biệt về mức độ phong phú tương đối trong CN đối với chuột CTRL ở cấp độ chi cho mỗi loài động vật. Chỉ các đơn vị phân loại có mức độ phong phú tương đối trung bình> 1% trong các nhóm này được hiển thị. Bản đồ nhiệt trên đỉnh của biểu đồ thanh cho thấy sự tăng / giảm của chi Akkermansia.

Xác định tác dụng phân tử của COLOSTRONONI đối với các phản ứng viêm toàn thân
Để tìm hiểu tác động của COLOSTRONONI đối với sự biểu hiện của cytokine ở chuột, chúng tôi đã đánh giá sự cảm ứng của các gen mã hóa IL-10, IL-12, IL-8 và TNF-α bằng cách sử dụng phương pháp qRT-PCR. Trong bối cảnh này, chúng tôi quan sát thấy biểu hiện cytokine khác nhau giữa nhóm chuột được bổ sung CN và nhóm CTRL [chuột được bổ sung dung dịch sucrose (2%)].
Kết quả thử nghiệm biểu hiện gen trên mô manh tràng chuột khỏe mạnh cho thấy mức độ biểu hiện IL-10, IL-12 và TNF-α giảm đáng kể có thể phát hiện được ở nhóm CN. Điều thú vị đặc biệt là đánh giá tỷ lệ giữa IL-10 và IL-12 và giữa TNF-α và IL-10.
Tỷ lệ IL-10 / IL-12 (khoảng 1: 1, trong điều kiện bình thường) [ 22 , 23 ], hiện đang được nghiên cứu như một chất chỉ điểm cho các bệnh viêm nhiễm và / hoặc chứng loạn khuẩn. Nó cũng hữu ích cho việc đánh giá tác dụng của việc bổ sung probiotics (tăng tỷ lệ do tăng biểu hiện IL-10) [ 23 , 24 ]. Sự giảm đồng thời IL-12 và IL-10 được nhấn mạnh (lần lượt là 74 và 78 lần) ( Hình 6 ) bảo toàn tỷ lệ IL-10 / IL-12 bình thường, phù hợp với việc thiếu những thay đổi trong thành phần hệ vi sinh vật trong hai thí nghiệm. các nhóm.

Mức ngang đại diện cho các trung gian. Trục y thể hiện mức độ biểu hiện dưới dạng biểu thức chuẩn hóa (ΔΔCt) theo phần mềm CFX96 Bio-Rad.
Tỷ lệ TNF-α / IL-10 giảm ở nhóm CN so với nhóm CTRL; kết quả này, do “đồng bằng” dương giữa TNF-α (-97 nếp gấp) và giảm biểu hiện gen IL-10 (23 nếp gấp có lợi cho IL-10) ( Hình 6 ) có thể được coi là dấu hiệu của một phản hoạt động viêm của COLOSTRONONI.
Điều thú vị là, COLSTRONONI dường như điều chỉnh phản ứng viêm bằng cách điều chỉnh tinh chỉnh biểu hiện gen TNF-α và IL-10 thay vì tạo ra sự điều hòa tăng IL-10. Việc giảm biểu hiện gen TNF-α, IL-10 và IL-12 đồng thời cho phép chúng tôi đưa ra giả thuyết rằng COLOSTRONONI ở đối tượng khỏe mạnh đóng một vai trò quan trọng trong cơ chế điều hòa miễn dịch thông qua việc giảm nhẹ tình trạng viêm sinh lý (mức TNF-α thấp và giảm TNF tỷ lệ -α / IL-10) duy trì, đồng thời, cơ chế sinh lý của sự dung nạp miễn dịch (duy trì tỷ lệ IL-10 / IL-12).
Tác động của COLOSTRONONI đến tính thấm của tế bào
Các thành phần chính trong nhóm xuyên màng bao gồm các phân tử claudins, occludin, và các phân tử kết dính liên kết (JAM) [ 25 ]. Một số dấu hiệu tính thấm của ruột này được đánh giá bằng qRT-PCR. Những dữ liệu này cho thấy không có sự khác biệt đáng kể nào trong biểu hiện gen đối với các claudin (claudin-1, claudin-2 và claudin-5), cũng như JAM-A và Occludin, ở những con chuột được điều trị bằng COLOSTRONONI (nhóm CN) so với những con không được điều trị chuột (nhóm CTRL). Những dữ liệu này cho thấy rằng việc bổ sung COLOSTRONONI không thể ảnh hưởng đến tính thấm ruột của những con chuột khỏe mạnh. Tuy nhiên, điều đáng nói là trong điều kiện khỏe mạnh, mức độ biểu hiện của gen TJs bình thường không bị sửa đổi.
Phần kết luận
Việc đánh giá các tính năng bổ sung tiềm năng của COLOSTRONONI trong điều kiện in vivo cho thấy khả năng của thực phẩm bổ sung này trong việc bảo tồn đỉnh cao hệ vi sinh vật đường ruột cũng như ảnh hưởng của nó đối với khả năng miễn dịch bẩm sinh của vật chủ có vú. Hơn nữa, phương pháp điều trị COLOSTRONONI đã kích thích sự gia tăng nhẹ của chi Akkermansia . Điều thú vị là Một . muciniphila là một loại vi khuẩn đường ruột, đã được đề xuất là một loài vi khuẩn tăng cường sức khỏe mới lạ do đặc tính điều hòa miễn dịch của nó.
COLOSTRONONI cũng cho thấy tác dụng làm giảm đáng kể sự biểu hiện của các gen mã hóa phản ứng IL-10, Il-12 và TNF-α, cho phép chúng tôi đưa ra giả thuyết về hoạt động điều hòa miễn dịch của thực phẩm chức năng này. Điều đáng chú ý là TNF-α làm trung gian cho những thay đổi trong biểu hiện của tế bào cốc và sự sulfo hóa mucin, và việc sản xuất nó có tương quan nghịch với tính thấm của biểu mô [ 26 , 27 ]. Điều thú vị là, việc giảm biểu hiện gen TNF-α trong nhóm CN lẽ cải thiện quá trình tổng hợp chất nhầy., Mà sau đó có thể thúc đẩy tăng nhẹ Một . muciniphila .
COLOSTRONONI không ảnh hưởng đến biểu hiện IL-8; kết quả này phù hợp với sự duy trì quan sát thấy tính toàn vẹn của biểu mô bình thường, được xác nhận bởi sự không điều chỉnh của các gen bao bọc cho claudin, tắc mạch và JAM, mà sự biểu hiện đã được chứng minh là được điều chỉnh theo cách phụ thuộc vào liều lượng bởi chính IL-8. [ 27 ]. Các kết quả này củng cố quan niệm rằng sữa non của bò và M . citrifolia ảnh hưởng đến khả năng miễn dịch bẩm sinh của vật chủ có vú.
Các nghiên cứu trong tương lai nên được thực hiện bằng cách sử dụng các lượng COLOSTRONONI khác nhau để đánh giá xem vai trò cân bằng nội môi của thực phẩm bổ sung này đối với hệ vi sinh vật đường ruột của động vật có vú có phụ thuộc vào liều lượng hay không.
Ngoài ra, sẽ rất thú vị khi đánh giá nếu vai trò nội môi của COLOSTRONONI đối với hệ vi sinh vật đường ruột cũng được khai thác trong các điều kiện viêm cấp tính như những gì xảy ra trong Bệnh viêm ruột (IBD). Trong bối cảnh này, một nghiên cứu gần đây liên quan đến mô hình chó IBD được điều trị bằng sữa non của bò (BC) đã chứng minh vai trò bảo vệ thú vị đối với hệ vi sinh vật đường ruột của động vật do BC điều khiển [ 28 ]. Cho đến nay, chúng tôi không biết liệu những tác động đã được xác định ở đây của COLOSTRONONI lên hệ vi sinh vật đường ruột ở chuột khỏe mạnh cũng có giá trị ở động vật có các triệu chứng IBD.
Trong bối cảnh này, các thử nghiệm lâm sàng in vivo trong tương lai sẽ có vai trò then chốt để khẳng định vai trò của COLOSTRONONI đối với thành phần hệ vi sinh vật đường ruột ở những đối tượng mắc bệnh viêm ruột (IBD), chẳng hạn như viêm loét đại tràng và bệnh Crohn.
Lời cảm ơn
Chúng tôi cảm ơn Guna Spa, Ý, đã cung cấp sản phẩm COLOSTRONONI. Nghiên cứu này được hưởng lợi từ cơ sở HPC (Máy tính Hiệu suất cao) của Đại học Parma, Ý.
Báo cáo tài trợ
GenProbio srl hỗ trợ Phòng thí nghiệm Probiogenomics. Nhà tài trợ cung cấp hỗ trợ dưới dạng tiền lương cho RA và GL và tài liệu nghiên cứu, nhưng không có bất kỳ vai trò bổ sung nào trong việc thiết kế nghiên cứu, thu thập và phân tích dữ liệu, quyết định xuất bản hoặc chuẩn bị bản thảo.
Dữ liệu sẵn có
Trình tự thô của cấu hình gen 16S rRNA có thể truy cập thông qua SRA ( https://www.ncbi.nlm.nih.gov/sra ), số gia nhập PRJNA497777.
Tài liệu tham khảo
1. Thuốc Cardani D. COLOSTRO NONI tác động lên sự đảo lộn của tế bào biểu mô, các sự kiện viêm nhiễm và tính toàn vẹn của hệ thống kết nối niêm mạc ruột . Minerva Gastroenterol Dietol . 2014; 60 ( 1 ): 71–8. Tập 2014/03/19.
2. Uruakpa FO, Ismond MAH, Akobundu ENT. Sữa non và lợi ích của nó: một đánh giá . Nutr Res . Năm 2002; 22 ( 6 ): 755–67. doi: Pii S0271-5317 (02) 00373-1 10.1016 / S0271-5317 (02) 00373-1
3. Nhà tài trợ JK, Steele JA, Schmidt DJ, Kim HB, Beamer G, Sun X, et al. Sữa non bò hyperimmune như một liệu pháp mới để chống lại nhiễm trùng Clostridium difficile . Tạp chí Bệnh truyền nhiễm . Năm 2015; 211 ( 8 ): 1334–41. Epub 2014/11/09.
4. Rathe M, Muller K, Sangild PT, Husby S. Các ứng dụng lâm sàng của liệu pháp sữa non bò: một đánh giá có hệ thống . Đánh giá dinh dưỡng . 2014; 72 ( 4 ): 237–54. Tập 2014/02/28.
5. Pakkanen R, Aalto J. Yếu tố tăng trưởng và yếu tố kháng khuẩn của sữa non bò . Int J sữa . Năm 1997; 7 ( 5 ): 285–97. 10.1016 / S0958-6946 (97) 00022-8
6. Abou Assi R, Darwis Y, Abdulbaqi IM, Khan AA, Vuanghao L, Laghari MH. Morinda citrifolia (Noni): Đánh giá toàn diện về việc sử dụng công nghiệp, hoạt động dược lý và thử nghiệm lâm sàng . J Chem Ả Rập . Năm 2017; 10 ( 5 ): 691–707. 10.1016 / j.arabjc.2015.06.018
7. Yang J, Paulino R, Janke-Stedronsky S, Abawi F. Hoạt động thu gom gốc tự do và tổng số phenol của nước ép và bột trái nhàu (Morinda citrifolia L.) trong chế biến và bảo quản . Thực phẩm Chem . Năm 2007; 102 ( 1 ): 302–8. 10.1016 / j.foodchem.2006.05.020
8. Zin ZM, Hamid AA, Osman A, Saari N. Hoạt động chống oxy hóa của các phân đoạn sắc ký thu được từ rễ, quả và lá của Mengkudu (Morinda citrifolia L.) . Thực phẩm Chem . Năm 2006; 94 ( 2 ): 169–78. 10.1016 / j.foodchem.2004.08.048 PubMed PMID: WOS: 000231989300001.
9. Furusawa E, Hirazumi A, Story S, Jensen J. Tiềm năng kháng u của một chất giàu polysaccharide từ nước quả của Morinda citrifolia (Noni) trên khối u cổ trướng sarcoma 180 ở chuột . Phytother Res . Năm 2003; 17 ( 10 ): 1158–64. Tập phim 2003/12/12.
10. Kamiya K, Tanaka Y, Endang H, Umar M, Satake T. Các thành phần hóa học của quả Morinda citrifolia ức chế quá trình oxy hóa lipoprotein mật độ thấp do đồng gây ra . J Thực phẩm Nông nghiệp Chem . Năm 2004; 52 ( 19 ): 5843–8. Epub 2004/09/16.
11. Saludes JP, Garson MJ, Franzblau SG, Aguinaldo AM. Các thành phần chống phân tử từ phần hexan của Morinda citrifolia Linn. (Họ Đào lộn hột) . Phytother Res . Năm 2002; 16 ( 7 ): 683–5. Tập phim 2002/11/01.
12. Groschwitz KR, Hogan SP. Chức năng hàng rào đường ruột: điều hòa phân tử và cơ chế bệnh sinh . Tạp chí dị ứng và miễn dịch học lâm sàng . Năm 2009; 124 ( 1 ): 3–20; câu đố 1–2. Tập phim 2009/06/30.
13. Komarova Y, Malik AB. Điều hòa tính thấm của nội mô qua con đường vận chuyển nội bào và xuyên tế bào . Annu Rev Physiol . Năm 2010; 72 : 463–93. Tập phim 2010/02/13.
14. Soini Y. Claudins trong các bệnh phổi . Phản hồi Res . 2011; 12 : 70 Tập 2011/05/31.
15. Ferrario C, Statello R, Carnevali L, Mancabelli L, Milani C, Mangifesta M, và cộng sự. Cách nuôi hệ vi sinh vật có vú trong ruột: Điều hòa vi khuẩn và trao đổi chất bằng chất xơ trong thức ăn . Biên giới trong vi sinh vật học . Năm 2017; 8 : 1749
16. Milani C, Hevia A, Foroni E, Duranti S, Turroni F, Lugli GA, et al. Đánh giá hệ vi sinh vật trong phân: một giao thức phân tích dựa trên gen torrent 16S rRNA ion được tối ưu hóa . PloS một . Năm 2013; 8 ( 7 ): e68739
17. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME cho phép phân tích dữ liệu giải trình tự cộng đồng thông lượng cao . Phương pháp bản chất . Năm 2010; 7 ( 5 ): 335–6.
18. Edgar RC. Tìm kiếm và nhóm các đơn hàng có độ lớn nhanh hơn BLAST . Tin sinh học . Năm 2010; 26 ( 19 ): 2460–1. Tập phim 2010/08/17.
19. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. Dự án cơ sở dữ liệu gen RNA ribosome SILVA: các công cụ dựa trên web và xử lý dữ liệu được cải tiến . Axit nucleic Res . Năm 2013; 41 (Sự cố cơ sở dữ liệu): D590–6. Tập 2012/11/30.
20. Lozupone C, Hamady M, Knight R. UniFrac — một công cụ trực tuyến để so sánh sự đa dạng của cộng đồng vi sinh vật trong bối cảnh phát sinh loài . Tin sinh học BMC . Năm 2006; 7 : 371 Epub 2006/08/09.
21. Derrien M, Belzer C, de Vos WM. Akkermansia muciniphila và vai trò của nó trong việc điều chỉnh các chức năng của vật chủ . Microb Pathog . Năm 2017; 106 : 171–81. Tập 2016/02/16.
22. Qiu X, Zhang M, Yang X, Hong N, Yu C. Faecalibacterium prausnitzii điều hòa các tế bào T điều hòa và các cytokine chống viêm trong điều trị viêm đại tràng do TNBS . J Crohns Viêm đại tràng . Năm 2013; 7 ( 11 ): e558–68. Tập 2013/05/07.
23. Gorska S, Sandstrom C, Wojas-Turek J, Rossowska J, Pajtasz-Piasecka E, Brzozowska E, et al. Sự khác biệt về cấu trúc và điều hòa miễn dịch giữa các exopolysaccharide lactobacilli được phân lập từ ruột của chuột bị bệnh viêm ruột gây ra trong thực nghiệm . Các báo cáo khoa học . 2016; 6 : 37613 Epub 2016/11/22.
24. Gad M, Ravn P, Soborg DA, Lund-Jensen K, Ouwehand AC, Jensen SS. Quy định trục IL-10 / IL-12 trong tế bào đuôi gai của người với vi khuẩn probiotic . Miễn dịch học FEMS và vi sinh y tế . 2011; 63 ( 1 ): 93–107. Tập 2011/06/29.
25. Campbell HK, Maiers JL, DeMali KA. Tác động qua lại giữa các mối nối chặt chẽ và nối các mối nối . Exp Cell Res . Năm 2017; 358 ( 1 ): 39–44. 10.1016 / j.yexcr.2017.03.061 PubMed PMID: WOS: 000407307100007.
26. Leppkes M, Roulis M, Neurath MF, Kollias G, Becker C. Chức năng Pleiotropic của TNF-alpha trong việc điều hòa phản ứng của biểu mô ruột đối với chứng viêm . Int Immunol . 2014; 26 ( 9 ): 509–15. Tập 2014/05/14. 10.1093 / Trim / dxu051.
27. Shen Y, Zhou M, Yan J, Gong Z, Xiao Y, Zhang C, et al. miR-200b ức chế bài tiết IL-8 do TNF-alpha gây ra và phá vỡ mối nối chặt chẽ của tế bào biểu mô ruột trong ống nghiệm . Am J Physiol Thuốc tiêu hóa gan Physiol . Năm 2017; 312 ( 2 ): G123 – G32. Epub 2016/12/17.
28. Filipescu IE, Leonardi L, Menchetti L, Guelfi G, Traina G, Casagrande-Proietti P, et al. Tác dụng phòng ngừa của việc bổ sung sữa non bò trong bệnh viêm đại tràng do TNBS ở chuột . PloS một . 2018; 13 ( 8 ): e0202929 Epub 2018/08/24.
